|
Ящеротазовые (Saurischia) — один из двух главных отрядов динозавров. Отличаются тем, что лобковые кости первично наклонены вперёд (позднее у некоторых представителей -- сегнозавры, дромеозавры и другие лобковые кости отклоняются назад).
|
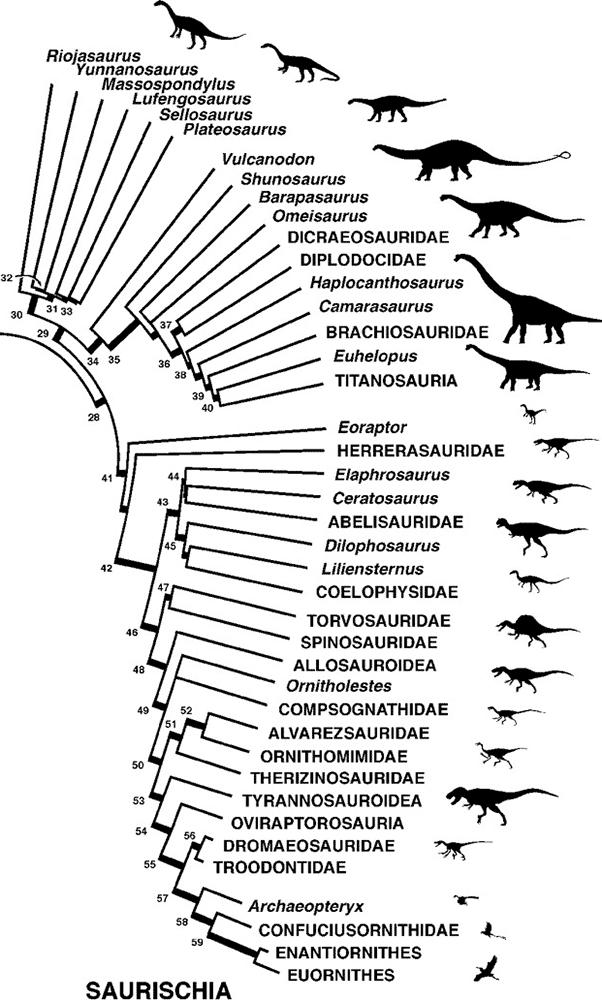
Таксономия подотряда Ящеротазовые
 |
| Бедренная кость тираннозавра - одного из ярких представителей группы ящеротазовых. |
 |
Ящеротазовых можно подразделить на две монофилетические группы (кладона) - тероподы и завроподоморфы.
Тероподы (звероногие) - это широкий спектр преимущественно двуногих форм, известных с позднего триаса до самого конца мелового периода. До недавнего времени их условно делили на две группы - целурозавров и карнозавров. Целурозавры - это двуногие животные мелких-средних размеров с длинными задними конечностями и маленькими, облегченными передними. Длинный хвост предположительно служил им балансиром при двуногой (бипедальной) локомоции. Находки показали, что целурозавры представляют собой не единый кладон, а несколько четко обособленных эволюционных линий. Долгое время их считали самыми примитивными тероподами, однако лидирует в этом смысле группа карнозавров, названная цератозаврами. Она представлена крупной юрской формой Ceratosaurus с небольшим рогом на носу, родом Coelophysis, известным по многим экземплярам из триаса штата Нью-Мексико, а также несколькими другими триасовыми родами из Северной Америки, Европы и Южной Африки.
Родственные связи различных групп целурозавров не вполне выяснены и по-разному интерпретируются специалистами. Особый интерес представляют их более эволюционно продвинутые формы, включая птиц. Среди них выделяются "страусоподобные динозавры", например Struthiomimus, Ornithomimus и Gallimimus. Это были бегающие тероподы средних-крупных размеров с облегченным телосложением. Передние конечности их длинные, но кисти не хватательные; задние ноги стройные, с компактными стопами; челюсти беззубые, морда тонкая и вытянутая. Только один род юрский, все прочие - меловые. Менее известны овирапторозавры, мелкие редкие двуногие формы длиной не более 2 м, также лишенные зубов. Однако их череп короткий и высокий, с сильно редуцированными лицевой частью и передними костями челюстей.
К наиболее интересным "некарнозаврам" относится группа т.н. дромеозаврид. Это двуногие существа мелких-средних размеров, известные из юры и мелового периода. Один из наиболее изученных их родов - Deinonychus из штата Монтана. Его четкий отличительный признак связан со строением стопы. Второй палец сильно увеличен, не служит опорой и оканчивается серповидным когтем, бывшим, вероятно, оружием нападения. Таким образом, стопа стала двупалой в противоположность обычной для динозавров трехпалости. Своеобразен также длинный и тонкий хвост, жесткий на протяжении примерно 3/4 своей длины, считая от конца.
К дромеозавридам близки редко встречающиеся троодонтиды, мелкие двуногие формы с самыми крупными среди всех динозавров относительными размерами мозга. Обе группы обладают многими признаками птиц.
Плотоядные карнозавры, например юрский Allosaurus и Tyrannosaurus из мелового периода, хорошо известны и представлены сравнительно немногими родами. Предшественником крупных юрских и меловых хищников был триасовый Palaeosaurus, довольно тяжелая двуногая плотоядная форма длиной ок. 2,7 м. В течение юрского периода доминировали гораздо более массивные ящеры, такие, как Allosaurus и Megalosaurus. Длина первого ок. 9 м. Его передние конечности сильно редуцированы, но еще достаточно крупные, чтобы функционировать. В противоположность ему у еще более крупных меловых форм, таких как Tyrannosaurus, передние конечности превратились в миниатюрные бездействующие придатки. Достигавший 15 м Tyrannosaurus был самым крупным когда-либо существовавшим наземным хищником. У этих гигантов голова относительно крупная. Вероятно, для снижения веса их череп приобрел крупные окна, не проиграв в прочности. Массивные челюсти усажены длинными, кинжалообразными зубами, прекрасно подходящими для разрывания жертвы. Похоже, что в течение юрского и мелового периодов эти хищники охотились на крупных растительноядных динозавров и, возможно, других рептилий соответствующих размеров. Главными их жертвами в юре были завроподы и стегозавры. В меловом периоде важным источником корма для этих крупных хищников, видимо, оставались некоторые завроподы, к которым добавились новые птицетазовые формы, в частности цератопсы и утконосые динозавры.
|
 |
| Кладистическая модель |
|
Завроподоморфы. В триасе появилась группа динозавров с такой явной тенденцией к увеличению размеров тела, что некоторые из ее юрских представителей стали самыми крупными из когда-либо обитавших на Земле сухопутных животных. Они передвигались в основном или полностью на четырех конечностях и были преимущественно растительноядными.
Прозавроподы. К этой группе относятся триасовые представители завроподоморф. Наиболее примитивен среди них род Thecodontosaurus из позднего триаса Англии, вероятно представляющий собой сестринскую группу по отношению ко всем остальным прозавроподам (т.е. параллельную им ветвь эволюции). Этот динозавр был двуногим. Прозавроподы в конце триасового периода быстро эволюционировали. Они известны на всех континентах, кроме Австралии и Антарктиды, и составляют неоднородную группу, включающую приблизительно 17 родов из 7 семейств. Типичным представителем может служить Plateosaurus, достигавший в длину около 6 м. Задняя часть его тела была очень массивной, как у многих теропод, однако довольно хорошо развитые передние конечности, вероятно, позволяли передвигаться как на двух, так и на четырех лапах. Широкие стопы мало походили на птичьи. Длинная шея несла маленькую голову. Сравнительно слабые челюсти усажены лопатовидными или листовидными зубами, которые напоминают известные у некоторых настоящих завропод.
Завроподы. Эти динозавры стали чемпионами по размерам среди наземных животных. Группа процветала в течение юрского периода и дожила, несколько потеряв в численности, до конца мела. Завроподы - гиганты среди сухопутных форм, однако их превзошли по величине некоторые современные киты, достигающие в длину 30 м. Apatosaurus (прежде известный как Brontosaurus) - массивное животное длиной ок. 21 м. Сравнительно стройный Diplodocus из юрского периода достигал в длину 24 м, включая очень длинный хвост. Brachiosaurus из Восточной Африки и Camarosaurus из Северной Америки были почти такими же длинными, но более тяжеловесными. Масса наиболее крупных из этих животных составляла, по существующим оценкам, ок. 50 т. Основная масса была сконцентрирована в тазовой области и передней части хвоста. Длинные, толстые, столбообразные задние ноги оканчивались стопами с подошвенной подушкой, как у слонов. Хвост довольно резко утончался в очень длинный гибкий "хлыст". Очень крупные туловищные и шейные позвонки несли глубокие боковые выемки, служившие для уменьшения веса. Передние конечности намного короче задних, хотя у Brachiosaurus и Camarosaurus и они довольно длинные.
Кладистическая модель 28 Saurischia; 29 Sauropodmorpha; 30 Prosauropoda; 31 Plateosauria; 32 Massospondylidae; 33 Plateosauridae; 34 Sauropoda; 35 Eusauropoda; 36 Neosauropoda; 37 Diplodocoidea; 38 Macronaria; 39 Titanosauriformes; 40 Somphospondyli; 41 Theropoda; 42 Neotheropoda; 43 Ceratosauria; 44 Ceratosauroidea; 45 Coelophysoidea; 46 Tetanurae; 47 Spinosauroidea; 48 Neotetanurae; 49 Coelurosauria; 50 Maniraptoriformes, 51 Ornithomimosauria; 52 Ornithomimoidea; 53 Tyrannoraptora; 54 Maniraptora; 55 Paraves; 56 Deinonychosauria; 57 Aves; 58 Ornithurae; 59 Ornithothoraces
У завропод были очень мелкие относительно общих размеров тела череп и головной мозг, но крупные глаза, а ноздри у некоторых видов располагались почти на макушке. Челюсти и зубы маленькие и слабые; вероятно, они обдирали ими, как гребенкой, мягкие части растений. Эти и другие особенности раньше считались адаптациями к жизни в воде, и завропод часто изображают стоящими в болотах или озерах. Однако, судя по ископаемым отпечаткам лап, эти гиганты передвигались и по сухому грунту. Анализ других признаков, например строения конечностей, также позволяет предположить, что главным местообитанием для них была твердая суша. Вертикальная ориентация мощных столбообразных ног и крайне длинная шея вполне могут служить приспособлениями к ощипыванию высоких деревьев. Наиболее вероятным источником корма для завропод были обильные в юрском и меловом периодах хвойные деревья.
Множество таких динозавров обитало в Северной Америке на протяжении всей юры. Характер осадочных пород и остатки растений и животных геологической формации Моррисон на западе США указывают на то, что ее отложения накапливались на дне широких долин, рек, ручьев, а также связанных с ними озер и болот. Растительность юрского периода представляла собой в основном вечнозеленые сообщества хвойных, семенных папоротников, саговников, гинкго и хвощей. Теплый климат, обилие зеленого корма и воды создавали прекрасные условия для жизни гигантских завропод.
Содержащие их остатки слои, сравнимые с формацией Моррисон, обнаружены также в юрских и нижнемеловых отложениях Европы и Восточной Африки. В течение мелового периода подходящие для завропод местообитания постепенно исчезали. Доминировать в растительности стали цветковые виды. Однако на протяжении значительной части мела завроподы еще продолжали существовать почти на всей Земле. В конце концов на границе между мезозоем и кайнозоем количество этих гигантов резко сократилось, и они быстро вымерли. В самых ранних кайнозойских отложениях, содержащих остатки типичных млекопитающих, завроподы не обнаружены.
|
|





 ICQ 470-479-001
ICQ 470-479-001
 Отряд
Отряд